Lipids/Fat: a turnkey nutrient for BSF larvae

When carbohydrates are abundant, BSF larvae transform them into lipids, storing them in their fat body through lipogenesis. This becomes particularly significant when their food has a low protein content. This lipid storage is crucial for adult fitness, as it generally boosts the reproductive capacity of flies. For instance, in locusts fed diets rich in carbohydrates but low in protein, there was an observed increase in lipogenesis (Zanotot et al., 2008)1, This is especially advantageous for females, as fat plays a vital role in egg development. Triacylglycerol, a type of energy-storing fat, makes up around 40% of a mature oocyte, which is a developing egg (Liu et al., 2017)2.
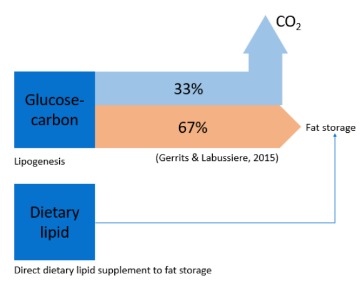
It’s worth noting that during lipogenesis, when carbohydrates like glucose are converted into fatty acids, only 4 out of every 6 carbon atoms are stored, while the remaining 2 are released as CO2 (Gerrits & Labussiere, 2015). However, it’s important to recognize that the process of depositing lipids carries significant physiological costs. Theoretical calculations indicate that the conversion of glucose into storage fat can consume as much as 20-25% of the energy provided by the food (Le Gall & Behmer, 2014)3.
Our understanding of the requirements for fat, whether in solid (fat) or liquid (oil) form, remains limited. As mentioned earlier, lipogenesis, as described by Equation 1 (taken from Frayn, 19834), necessitates 83 units of glucose to produce 6 units of fat storage. This process results in a 33% loss due to carbon atoms being emitted as CO2. This raises the question: why not directly provide dietary lipids that can contribute to larval fat storage?
83 ???????+30 ?2−6 ???????????????+169 ??2 (1)
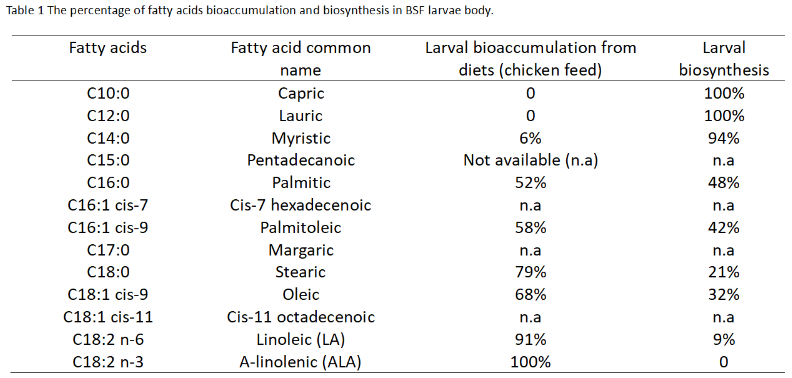
To address this question, we must first identify the specific fatty acids that BSF larvae can accumulate. Thanks to the team of Hoc et al. (2020)5, sheds light on by tracing hydrogen isotopes to understand fatty acid biosynthesis in BSF. Larval body fat is composed of fatty acids either synthesized within their bodies or acquired from their diet.
For instance, palmitic acids, a major component found in oil palms, make up 58% of the larval palmitic acid content, which is primarily obtained from their standard diet (chicken feed). Similarly, stearic and oleic acids, abundant in animal fat, can be directly assimilated from the larval food and contribute to 60-80% of the stored fat (see Table 1). However, BSF larvae lack the ability to synthesize polyunsaturated fatty acids (PUFA), and PUFA may be essential for their development. For example, C18:2 n-3 (omega-6) cannot be synthesized by BSF larvae and is entirely acquired from their diet, accounting for 100% of its presence.
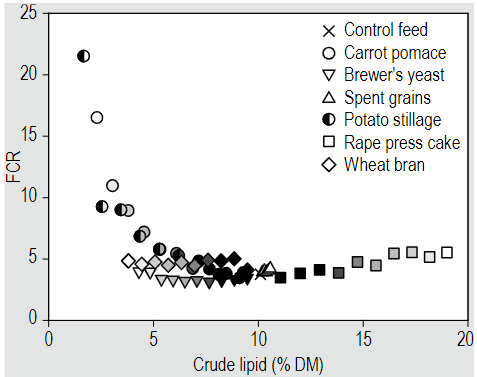
In a recent publication by M. KieBling et al. (2023)6, they observed that the Feed Conversion Rate (FCR), where a lower value indicates higher efficiency, significantly increased when the crude lipid content in the substrate was below 4%, regardless of the substrate type. Reduced larval performance in these instances can be attributed to a deficiency in essential fatty acids crucial for larval development. Conversely, high lipid contents (>15%) could result in excessive free fat in the substrate, creating anaerobic and sticky conditions on the larval surface. This disrupts larvae respiration and hinders their growth. Consequently, M. KieBling et al. (2023)9 concluded that a crude lipid content of about 10% on a dry matter basis in the substrate could be optimal for achieving a low FCR.
- https://doi.org/10.1111/j.1365-3032.1993.tb00617.x
- https://doi.org/10.1371/journal.pone.0182601
- https://doi.org/10.1093/icb/icu102
- https://doi.org/10.1152/jappl.1983.55.2.628
- https://doi.org/10.1038/s41598-020-68784-8
- https://doi.org/10.3920/jiff2022.0096
Comments on this post
Comment posted by Oscar Julian Arroyave S:
Kind regards from Colombia, although it is true, the larvae degrade any organic residue, if you want to obtain good results, you must balance the diet taking into account the protein/CNS, FB, fat and moisture ratio and it is difficult to find an organic substrate that meets all this on its own.
Comment posted by Ssimbwa David Jograss:
I live in rural area, there is cured where banana and mangoes are squeezed for their juice and packed.
I collect the resulting organic waste and feed the larvae of Black Soldier Fly.
I notice some challenges.
1. With banana waste maggots are very big in just seven days.
2.With mangoes waste larvae take 14 days to attain half size of 1. above!
In both cases my free range birds feed very well.
Thus in a kg of larvae in 1 I get 30kg in 21days, while in 2. I get far less.










